Bài 12B. Kiểm soát chu kỳ tế bào
Các phân tử kiểm soát CKTB
Nhóm protein thúc đẩy phân bào
Protein Cdk:
- Là các kinase phụ thuộc cyclin. Mỗi Cdk điều hòa hoạt động tại một điểm k.soát khác nhau trong CKTB: CDK4 ở đầu G1; Cdk2 ở cuối G1 và đầu S; Cdk1 ở G2
- Hàm lượng tương đối cao và ổn định nhưng đều ở dạng bất hoạt
- Đều có hoạt tính kinase, gây phosphoryl hóa, khi l.kết cyclin tương ứng
Protein cyclin:
- 4 loại: G1-cyclin (cyclinD); G1/S-cyclin (cyclinE); S-cyclin (cyclinA) và M-cyclin (cyclinB),
- Tín hiệu nội - ngoại bào và thời gian của CKTB
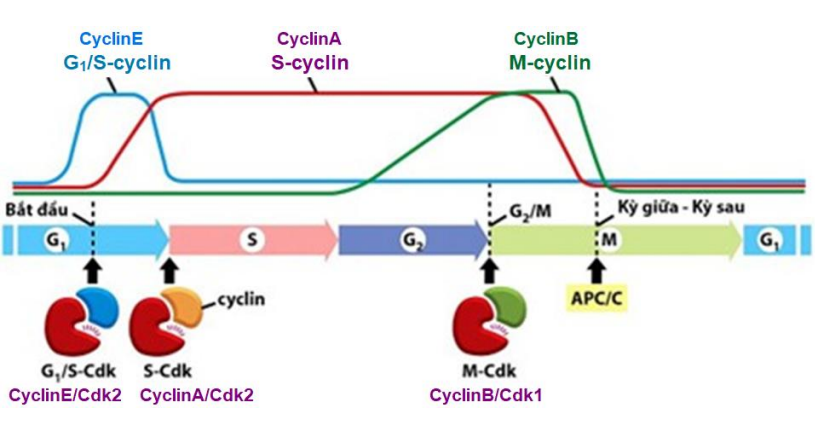
Khi tb chuyển sang gđ tiếp theo hoặc đã vượt qua từng điểm k.soát, cyclin nào đã hoạt động trước đó đều bị thoái hóa bởi các enzyme bào tương
Nhóm protein gây ức chế, làm dừng CKTB
Ức chế hoạt tính kinase của các Cdk
- Protein ức chế Cdk (CKIs): p16, p19, p21 và p27 * Vai trò: kiểm soát quá trình phân bào và ức chế hiện tượng tăng sinh u
- Hoạt động: Khi tế bào gặp yếu tố bất lợi, CKIs gắn trực tiếp vào các Cdk
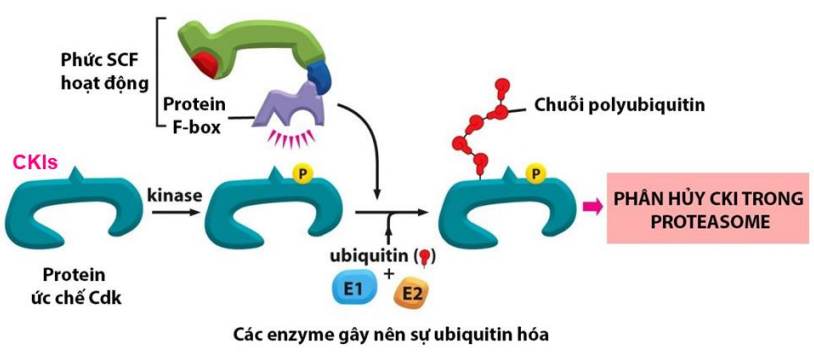
Khi xong nhiệm vụ, CKIs bị thoái hóa do phức SCF và protein F-box hoạt động (c.đường ubiquitin-proteasome) * Phức hợp SCF chỉ hoạt động khi các CKI đã được gây phosphoryl hóa trước đó.
Gen p16 (9q21)
Tb bị stress
- ĐBG p16
Cơ chế phân tử tại 3 điểm kiểm soát
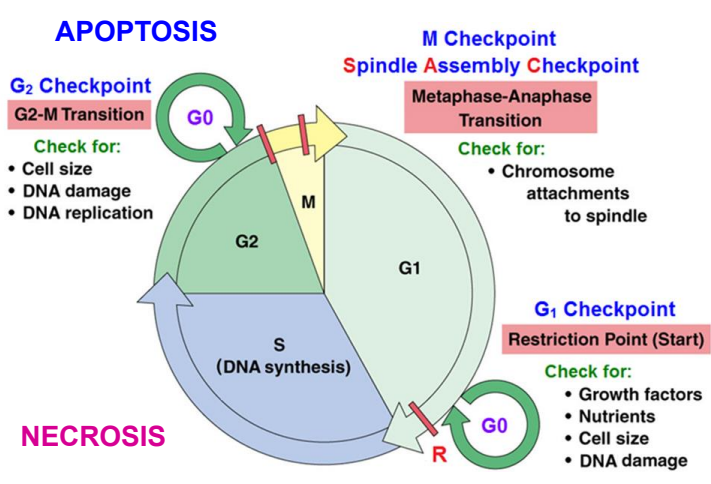
Điểm k.soát cuối G1 (điểm R, bắt đầu)
Là điểm k.soát mà tb cam kết không được đảo ngược q.trình phân bào khi đã vượt qua
Cần phải có: GF - Kích thước đủ lớn - Chất dự trữ - DNA toàn vẹn
Nếu điểm R phát hiện bất thường trong tb
Cơ chế:
- Khi GF tác động tb ở đầu G
- G1/S-cyclin hoạt hóa Cdk2
- Đầu gđ S, lượng S-cyclin tăng lên và duy trì đến hết KG
⟹ Khởi động sao chép DNA (rất cần sự hiện diện của G1/S-cyclin và S-cyclin)
Điểm k.soát G2/M
Đảm bảo các yêu cầu ở điểm k.soát cuối G1 * các sợi nhiễm sắc phải được sao chép 1 lần và không bị lỗi.
Khi điểm k.soát G2/M phát hiện bất thường trên DNA
Vào đầu G2, M-cyclin được tạo ra, bắt đầu tăng cao và tồn tại đến khi tb vào kỳ sau.
- M-cyclin hoạt hóa Cdk1
- Các gen ngừng phiên mã
- Vỏ nhân tan rã
- Thoi phân bào thành lập
- Co ngắn sợi nhiễm sắc
- Cuối thời kỳ M, M-cyclin bắt đầu giảm, G -cyclin lại 1 được tổng hợp tăng lên để tiếp tục CKTB tiếp theo.
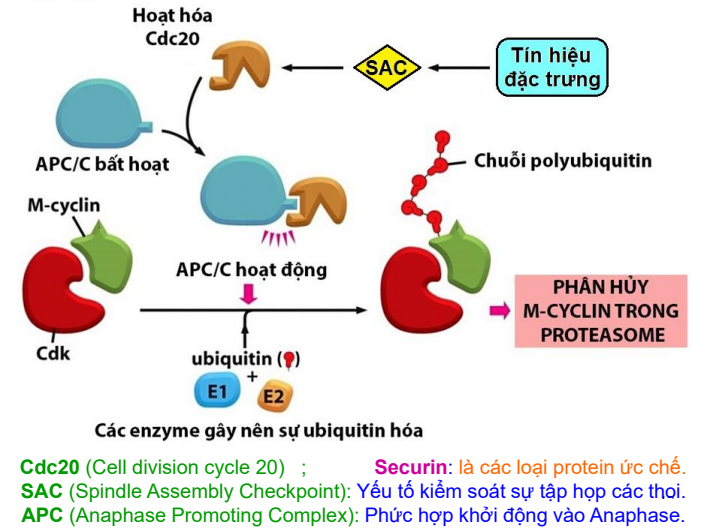
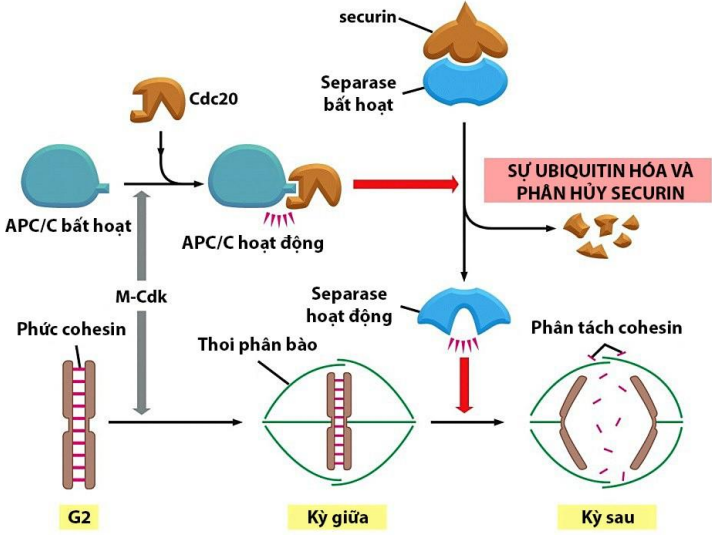
Điểm k.soát M (ở cuối KG và đầu KS)
Ngoài M-Cdk, các phức hợp khác cũng xuất hiện ở KG: SAC, APC/C, Cdc20
Cơ chế:
- Khi OVT tâm động gắn lên tâm động của các NST kép
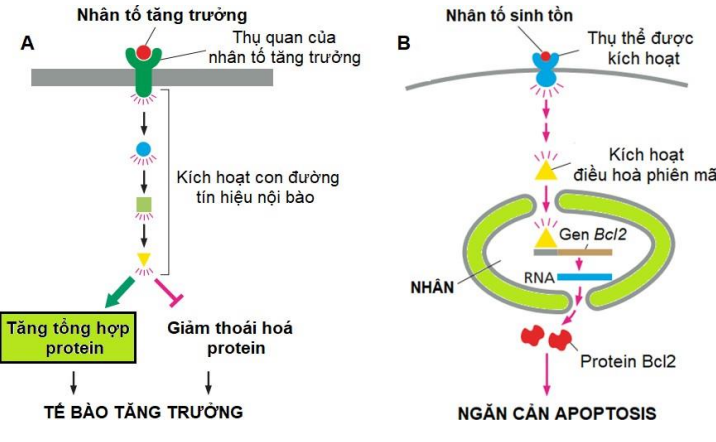
Điều hoà bởi các yếu tố ngoại bào
Các nhân tố ngoại bào (chủ yếu là các protein tín hiệu được tiết ra từ các tb lân cận,…).
- Các nhân tố ngoại bào thường l.kết các thụ thể trên bề mặt tb
Các nhân tố tăng trưởng kích thích tb phát triển
(tăng kích thước và khối lượng tb)
- Các nhân tố Gf thúc đẩy tb tăng tổng hợp protein và ức chế các protein và đại phân tử khác phân hủy.
- Cơ thể muốn phát triển lệ thuộc: sự phát triển và sự phân bào. Nếu: tế bào có phân chia mà không phát triển
- Ở nấm men, sự phát triển và phân bào đều cần chất dinh dưỡng. Ở tb ĐV đều cần tín hiệu từ các tb khác.
- Tb phát triển không lệ thuộc hệ thống k.soát CKTB, ngược với phân bào (hiểu biết khá rõ). Vd: tb TK, Cơ.
Các nhân tố sinh tồn (Sf) ngăn cản quá trình apoptosis, thúc đẩy tb tồn tại
- Mỗi tb đều cần nhân tố sinh tồn từ các tb khác để tồn tại (do tăng hoạt động nhóm anti-apoptotic Bcl-2). Nếu thiếu, tb tự kích hoạt apoptosis. Vd: Các tb TK
- Các tb TK thường được tạo ra quá nhiều vào gđ sớm ở cơ thể đang phát triển
Các nhân tố gây phân bào (M) bằng cách thúc đẩy tế bào vào S
Tb cần tăng sinh phải có nhân tố M: PDGF, TGF-α, EGF,… đều có tính đặc hiệu rộng, trừ erythropoietin.
Khi liên kết với thụ thể
- Tăng phiên mã các gen
- Hoạt hoá G -cyclin
Có nh.tố M
- PDGF là protein tiết có trong các túi tiết của tiểu cầu. Khi cục máu đông hình thành tại vết thương, PDGF (được giải phóng từ các tiểu cầu trong cục máu đông) l.kết thụ thể Tyrosine kinase của các tb còn sống xung quanh
- Nếu 1 phần mô gan bị mất (phẫu thuật,…), PDGF ở cục máu đông kích thích các tb gan còn sống xung quanh đó tăng sinh
Hầu hết các tb người đều ở G0 , nhưng khả năng đảo ngược gđ G0 thì khác nhau giữa các loại tb. Vd:
- Các tb TK và tb cơ xương (tb biệt hóa cuối cùng) đều ở G0 mà gen mã hóa Cdk và cyclin luôn bị tắt vĩnh viễn
- Các tb gan khi ở G0 , nếu bị tổn thương
- Nguyên bào sợi và một số B.c lymphocytes có thể ra vào CKTB nhiều lần trong quá trình sống.
Một số nhân tố ngoại bào ức chế phân bào
Các nhân tố này ức chế phân bào thay vì thúc đẩy như 3 loại nhân tố kể trên. Vd:
- TGF-β ức chế 1 số loại tb tăng sinh bằng cách ngăn cản sự tiếp diễn CKTB ở G1 hoặc kích thích apoptosis
- Myostatin là protein ức chế các tb cơ tiền thân (nguyên bào cơ) phát triển và tăng sinh.
- Ở động vật, nếu thiếu myostatin hoặc được điều trị bằng các chất ức chế myostatin